Significance of the Bluefish Caves in Beringian Prehistory – Page 1
THE SIGNIFICANCE OF THE BLUEFISH CAVES IN BERINGIAN PREHISTORY
Jacques Cinq-Mars
Curator, Quebec Archaeology
Archaeological Survey of Canada
Canadian Museum of Civilization
FOREWORD
This is a translation of a paper originally published, in French, in Revista de Arqueología Americana, No. 1, (1990): pp. 9-32. But for a few minor corrections and the presentation of upgraded illustrations (Fig. 1- 5) and faunal list (Table 1), this Web version is essentially the same as the original one.
The 1989 article by Cinq-Mars and Morlan (referred to in the text as “Cinq-Mars and Morlan 1989”) was finally published as:
Cinq-Mars, Jacques and Richard E. Morlan. 1999. “Bluefish Caves and Old Crow Basin: A New Rapport,” in Ice Age Peoples of North America, ed. by Robson Bonnichsen and Karen L. Turnmire, pp. 200-212. Corvallis: Oregon State University Press for the Center for the Study of the First Americans.
On the other hand, the article by Cinq-Mars and Nelson (1989 — also referred to in the text) has not as yet appeared.
For additional details on the available 14C dates from Bluefish (briefly mentioned in the text), see the Canadian Archaeological Radiocarbon Database (CARD) at:
www.canadianarchaeology.com/radiocarbon/card/card.htm
Jacques Cinq-Mars, Hull, February, 2001
INTRODUCTION
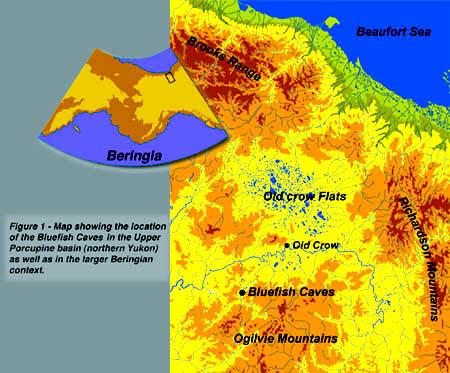
About 400 years ago, responding to his era’s need to explain the Aboriginal presence in the New World, José de Acosta postulated what would become known as the Bering Strait, i.e., a place where the relative proximity of Asia and America would have allowed populations to move between the two continents. Today, this pathway and the adjacent regions (Fig.1) still provide the geographical canvas on which numerous more or less viable hypotheses have been woven, variations on the theme of the peopling of the Americas. Following many recent studies, and in an attempt at biogeographical synthesis, these regions have come to be known as Beringia. (Hopkins 1959, 1967; Hopkins et al. 1982; Hultén 1968). For our purposes we shall refer more precisely to eastern Beringia, the territory corresponding to those parts of Alaska and the Yukon that, with the exception of several alpine zones, were spared the glacial advances of the Pleistocene.
Spurred on by continuing archaeological questions, interdisciplinary research in the region over the past few decades has enabled us to form a clearer picture of Beringian palaeogeography. This research has also clarified various aspects of the evolution of the Pleistocene environment to which human populations had to adapt, some of them eventually spreading south. The Beringia concept, and its variations and derivations (Note 1) , are essential components of any archaeological discussion treating the origins of the first Aboriginal populations in the New World within a Pleistocene time frame. It must be noted, however, that relatively little pertinent data have come out of these investigations, mainly because of the difficulties inherent in archaeological research in these regions. It seems appropriate, therefore, to clarify several points of Beringian archaeology that have often been “hot topics,” at least in certain hemispheric contexts.
One can divide the archaeological findings, as well as their most recent and plausible interpretations, into two groups. The first group pertains mainly to Late Glacial times, and comprises a complex set of data from a growing number of sites discovered in Alaska and the Yukon. Without being conclusive, this information seems more and more likely to provide answers to the question of the current peopling of the boreal and arctic regions of the Northwest, as well as that of Clovis origins. In the latter case, the findings tend to satisfy supporters of a “short-chronology”. The second set of data is not as telling, and at present does not appear to be continuous with the first one. It is mainly the result of research carried out in the northern Yukon, and whose findings suggest older cultural manifestations, dating back to the Glacial Maximum or even earlier, as we shall see. These findings provide support for the “long-chronology” school of thought.
By examining some of this material, and more particularly, evidence from the Bluefish Caves in northern Yukon, we shall attempt to shed a Beringian light on several facets of this origins-centred archaeology.
Significance of the Bluefish Caves in Beringian Prehistory – Page 2
DESCRIPTION OF THE BLUEFISH CAVES
Location and environment
The Bluefish Caves (Note 2) are located 54 km southwest of the Vuntut Gwichin village of Old Crow, overlooking the middle course of the Bluefish River, a tributary of the Porcupine River (Fig.1). This is a region of Devonian limestone hills (Norris 1985) at the northern end of the Keele Range, which in turn forms the foothills of the great massif of the Ogilvie Mountains that mark the centre of the Yukon. These hills also mark the southwestern edge of the enormous network of lacustrine basins, which during the upper Pleistocene were inundated by the waters of the Bluefish, Old Crow and Bell glacial lakes (Hughes 1972). The environment is characteristic of boreal forest in mountainous regions (Ritchie et al., 1982), with spruce trees (Picea glauca and Picea mariana) on the pediments and more or less continuous tundra zones along ridges over 750 m high. The climate is typical of this boreal type of environment, and the landscape has been shaped by multiple periglacial processes characteristic of continuous permafrost regions.
Geomorphological context

Map of the Bluefish Caves.
The caves are found at the western extremity of a ridge dominating a narrowing of the Bluefish River. They are nestled at the foot of a series of limestone outcrops standing about 250 m above the river valley. There are three caves (I, II and III), actually small cavities whose volume ranges from about 10 m3 to 30 m3 (Fig. 2). Contrary to what had been suggested earlier (Cinq-Mars 1979), they appear to have been formed in a very broad geographical and chronological context namely, that of a regional karst landscape now under study (Cinq-Mars and Lauriol 1985; Lauriol et al. 1989; Roberge et al. 1986). In other words, these are not cavities formed by congelifraction, but rather the remains of a former, greatly reduced karst network uncovered by the erosion of the slopes.
Deposit
Sampling and excavations both inside and outside the three caves, as well as subsequent sedimentary analyses, have revealed a depositional sequence characterised by a relative uniformity both in terms of the sedimentary sequence and the mode of deposition (Fig. 3). This sequence can be summarised as follows:
- The floor is a rocky surface (bedrock) studded with cryoclastic fragments, with some residual areas of highly altered sediments in fissures and depressions.
- The overlying sediment, of eolian origin, is relatively homogeneous loess. It may reach more than a metre deep and contains variable quantities of cryoclastic elements, mainly from the walls and ceiling of the cavities and from the cliff faces which rise about 10 m over the entrances.
- Results of granulometric and sedimentological analyses on samples collected under the dripline of Cave II suggest that, despite its apparent homogeneity, the loess can be divided into three facies which reflect both the transport conditions of the sediments and their place of origin (Note 3)
- Overlying the loess, gradually or discomformably, is a layer of humus-rich cryoclastic rubble which, outside of the caves, can be slightly more than a metre deep. The thickness of this unit declines rapidly at the cave entrances where it becomes, on the interior of the cavities, a simple organic enrichment of the upper portion of the loess.
- Finally, the surface of the fill is characterised by herbaceous and shrubby vegetation on the exterior of the caves, and by a discontinuous cover of ferns, mosses and lichens on the interior.

Bluefish Cave II
action of periglacial phenomena such as cryoturbation and congelifluction, and which can sometimes make interpretation difficult. These deposits do not lend themselves readily to a precise stratigraphic reading, and more than in most locations, decoding them requires the contribution of other types of data.
Palynological data
The analysis of sediments from the interior of Cave I and the exterior of Cave II has provided pollen diagrams which, despite the nature of the deposits, are indicative of a certain degree of depositional integrity. These diagrams have already been published (Cinq-Mars 1979; Ritchie et al. 1982) and so we will outline only the most significant details here:
- First, corresponding stratigraphically to the lower level of the loess, we find a pollen assemblage presenting the characteristics of tundra rich in herbaceous species.
- Above this, in the upper level of the loess, is a zone characterised mainly by an increase in, and predominance of birch (Betula).
- Finally, in the portion of the deposit which has been described as humus-rich rubble, a third assemblage is characterised by a discernible decrease of herbaceous species and a significant increase in spruce (Picea) and alder (Alnus).
This pollen biostratigraphy, though only weakly defined, holds up well in relation to the findings of other palynological research carried out in the region (Ritchie and Cwynar 1982; Ritchie 1984). It is therefore apparent that the lower portion of the loess contains traces of a xeric herbaceous tundra environment. Following these environmental conditions, which differ vastly from what we find today, there is a series of changes, starting with the development of shrubby birch tundra and leading eventually to the rapid establishment of boreal forest conditions quite similar to the ones we see at present time.
This correspondence provides several useful chronological markers. It would appear that the development of shrubby tundra, with increasing birch, dates to around 14,000-13,500 BP; and that conditions suitable for the development of boreal spruce forest in the region appeared around 10,000 BP. Finally, it must be emphasised that the correspondence between the Bluefish pollen diagrams and the regional ones supports the idea noted above of the relative integrity of the deposits, and this in spite of the inevitable periglacial scrambling.
Palaeontological data
All three caves have yielded thousands of bone remains, which are extremely well preserved due to the very favourable sedimentological and taphonomic conditions. The fauna comprises both large and small mammals, including a significant series of microtines, as well as birds and fish (Table 1). It can be divided into two series, found in the loess and in the humus-rich rubble respectively.
The loess contains a wealth of fauna, in terms of quantity, complexity and diversity, which correspond to Guthrie’s (1982, 1985) description of Beringian late Pleistocene mammoth steppe fauna. The megafauna includes horse (Equus lambei), caribou or reindeer(Rangifer tarandus), sheep (Ovis dalli), bison (Bison priscus), moose (cf. Alces alces), wapiti or elk (Cervus elaphus) and mammoth (Mammuthus primigenius). There are also saiga (Saiga tatarica), muskox (Ovibos moschatus), bear (Ursus), wolf (Canis lupus) and lion (Panthera).
In contrast, both quantitatively and qualitatively, the bone remains found in the humus-rich rubble show characteristic signs of a significant impoverishment of the regional megafauna, undeniable indication of mass extinction and extirpation on a regional and continental scale. In fact, this unit yielded only a few sparse specimens of caribou and sheep, a faint echo of regional Holocene megafauna that also includes moose, bear and wolf.
This contrast in the fauna, like that seen in the palynological record, is concordant with a definite chronostratigraphic break (loess/humus-rich rubble) (Note 4). Its significance, which remains to be fully understood, seems to be related to global ecological changes in Beringia and many other parts of the world during the last millennia of the Late Glacial (Lundelius 1989).
Chronological context
As we have seen, the sedimentological, palynological and palaeontological data enable us to place the cave deposits on a chronological scale that definitely includes the Holocene (humus-rich rubble) and the end of the late Pleistocene, or Late Glacial (loess). Several 14C measurements made early in the research (Cinq-Mars 1979; Morlan and Cinq-Mars 1982) roughly confirmed this chronostratigraphic estimate and even gave some indication of precision. We were able to date an episode of forest fire, definitely Holocene, in the (cryoturbated) upper sediments of Cave I. We also obtained a date of 12,900 BP from the femur of a horse, collected in the upper level of the lower loess of Cave I; and a date of 15,500 BP from a mammoth scapula found in the lower loess of Cave II, where the palynological signs of the herbaceous tundra were identified.
A new series of 14C dates obtained recently from specimens of megafauna (Note 5; Cinq-Mars and Nelson 1989) recovered from the three caves have enabled us to put forward a somewhat more detailed chronostratigraphy which sheds new light on a number of palaeontological, palaeoecological and archaeological issues.
For example, some of these findings can help us identify periods of extinction and extirpation and, in retrospect, furnish valuable information on the evolution of the environment. The example of the saiga is particularly instructive. Its ecological requirements (Vereshchagin and Baryshnikov 1982) make it an unusually fine indicator of a xeric environment (semi-arid steppe), relatively rich in herbaceous species and characterised by only slight accumulations of snow in winter. Until recently, the presence of saiga in eastern Beringia was dated prior to the Glacial Maximum (Harington 1980; Matthews 1982). However, its discovery in the sediments of Cave III indicate that populations of saiga, apparently living in an environment which was not altogether unfavourable, survived in far-eastern Beringia as late as about 13,400 BP. The overlap between this date and the age of the (palynological) birch zone which, as we have seen, is an indicator of a major ecological change, probably means that the Bluefish saiga is one of the last to have lived in these regions.
Besides providing clear confirmation of the loess/humus-rich rubble dichotomy and of its dating to about 10,000 years ago, many of these new chronometric results have led to an unexpected increase in the range of our chronostratigraphy. As we shall see below, some results indicate that the history of the deposit goes as far back as about 25,000 BP. More importantly, these results demonstrate that the mammoth steppe fauna mentioned above, in a combination that remains to be precisely determined, constituted an essential element of the Glacial Maximum biotope of eastern Beringia (between about 17,000 and 25,000 BP). Without elaborating further on this information, which sheds light on an important Beringian controversy (Note 6), we must emphasise that this demonstration of the viability of a Glacial Maximum environment is of major significance for our understanding of several aspects of Beringian archaeology.
- First, corresponding stratigraphically to the lower level of the loess, we find a pollen assemblage presenting the characteristics of tundra rich in herbaceous species.
Significance of the Bluefish Caves in Beringian Prehistory – Page 3
CULTURAL MANIFESTATIONS
Like other Beringian sites that have yielded very ancient archaeological material, the cultural remains found in the Bluefish Caves are exasperatingly sparse. They consist of three classes of lithics, some butchering marks found on various megafaunal remains and some examples of simple bone tools. With a few rare exceptions (Note 7) , these were all found in the loess layer of Caves I and II, and in a context which, once again, is not always easy to read.
Lithic artefacts
The first class consists of a series of about one hundred specimens. It includes, on the one hand, shaped objects (Fig. 4) such as cores, microblades, angle burins on truncated blades or flakes, burin spalls, notches, etc. and, on the other hand, various flakes and flake fragments, some of which may have been used. The raw materials are primarily high-quality cherts, usually blue, but occasionally speckled or more rarely, black, and definitely exotic to the region, although we have not yet determined their exact source (Note 8). So far, such artefacts have been recovered only in Caves I and II.

Most of them occurred mainly in the interior and at the entrance of Cave II, both at the bottom and the upper limit of the (lower) loess level where were also found most of the faunal remains. Several technological characteristics of the burins (all angle burins on truncated supports) may indicate that much of the assemblage represents either a specific episode in the use of the cave, or consecutive visits by a particular group. Although the context does not allow us to date these lithics with precision, we know that they considerably predate the end of the period in which the loess was deposited; i.e., before 10,000 BP. Their provenance and association with the remains of extinct species suggest that they may have been incorporated into the deposit around 12,000 BP or perhaps even earlier.
Several tools, including a burin spall, a few microblade fragments and some flakes (one resulting from bifacial retouch) were also collected inside Cave I, in a somewhat more precise stratigraphic context (Cinq-Mars 1979). Some of these pieces seem to date from the first half of the period characterised by the “birch” zone. Others are slightly more recent (possibly between 10,000 and 12,000 BP) or earlier (before 13,500 BP).
The second class consists of several dozen detrital microflakes, the result of flaking, retouching or using chert tools (Note 9). Small as they are, they exhibit all the morphological characteristics of larger flakes or splinters, and are found, in varying quantities, in any archaeological deposit where stone tools were made or used. The raw materials they are made of differ significantly from that of other stone fragments of similar sizes. Although microflakes were found in all three caves, those from Cave I furnished the most information. Their distribution inside the cave parallels that of the tools described above and is also suggestive of some cultural sedimentation in that portion of the loess unit whose pollen is characteristic of the herbaceous tundra which dates, as noted above, from between 25,000 and 13,500 BP.
The third class comprises a number of small cobbles, which for the most part were collected at the base of the loess, at or near the bedrock, and generally near the entrance or at the front of the caves. As they are all clearly allochthonous, we believed until recently (Morlan and Cinq-Mars 1982: Fig. 9) that their presence in the loess could be explained only by animal activity or in the case of the larger specimens, possibly by cultural activity. However, as is the case with the caves themselves, their origin is best explained in the context of the development of the regional karst network. Nevertheless, we have chosen to retain this category, as it may eventually be of use for several of the larger specimens that could, after all, have been used as hammerstones
Cut or butchering marks
Despite the difficulties inherent in any attempt at taphonomic interpretation, we have managed to extract complementary signs of human presence from the enormous palimpsest (Binford 1981: 9) represented by faunal and other remains from Bluefish.
The evidence consists of a variety of cut marks, incisions, scrape marks, chopping marks and striations resulting from the intentional butchering and defleshing of animals with stone tools, and penetrating, more or less deeply and in various places, the external walls of the bones. (Morlan and Cinq-Mars 1982: Fig. 10). It is important to note that we refer here to undeniable cultural indicators and not to similar marks made by carnivores, rodents, various geological processes or even excavators (Note 10). Thus far, we believe that we have been able to identify examples on numerous elements of the skeleton of nearly every large mammal species, with the possible exception of wolf, moose, wapiti and saiga. Almost all were found in Caves I and II.
This type of data also enables us to refine the time frame of the cultural content of the deposit. As there are no such markings on the faunal remains found in the humus-rich rubble, it is evident that this type of evidence and its causes date to the Pleistocene. This is confirmed by the 14C dates mentioned earlier and which were obtained from some of the specimens exhibiting such modifications. These dates suggest that cultural activities relating to the exploitation of the Bluefish fauna occurred sporadically between about 25,000 BP and 10,000 BP.
Bone toolmaking
The few examples of bone toolmaking can be divided into two groups. The first includes traditional tools that have been shaped somewhat, either intentionally or through use. Examples were collected in Caves I and II. The second group includes bones that were shaped by percussion. There are fewer examples of these, all from Cave II.
The first group comprises a few long bones which, after having been split lengthwise, may have been used as fleshers for processing hides. One in particular, shaped from a caribou tibia, exhibits a planed facet which may have been made with a burin, as well as a highly polished area located along the edge of a distal break (possibly the result of use ?) (Morlan and Cinq-Mars 1982: Fig.9). This object was discovered outside Cave II, in the lower level of the loess. It has been dated to 24,820 BP, giving us a clearer picture of the chronological range not only of the deposit, but also of its cultural manifestations. In other words, we believe that we can add this tool to a growing list of data which, while sparse, demonstrates that human populations were in a position to exploit the resources of the region during the Glacial Maximum or even earlier.
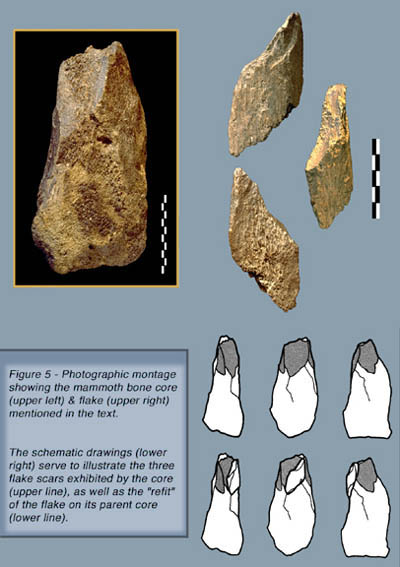
The same goes for the second group that comprises only two bone objects produced by percussion. They consist of a bone flake (Note 11) and its parent core (Fig. 5), both obviously derived from the same mammoth long bone. They were obtained through a relatively complex sequence of actions or “chaîne opératoire”, which can be summarised as follows:
- the raw material, namely a mammoth long bone, was first reduced to a fragment consisting of an epiphysis and the contiguous portion of the diaphysis;
- what could be described as a rough striking platform was then prepared at the end of the diaphysis segment;
- from this platform, a series of three flakes, ranging from 7 to 10 cm in length, were subsequently detached by percussion from the cortical face of the diaphysis segment;
- finally, one of these flakes, the longest one, was further worked and/or retouched bifacially and reduced diagonally, from its proximal end, by more than a third of its original size.
Significance of the Bluefish Caves in Beringian Prehistory – Page 4
INTERPRETATION AND CONCLUSIONS
Admittedly cryptic and fragmented, these bits of information do, nevertheless, provide us with a coherent picture of some of the palaeoecological and cultural phenomena that characterised the eastern confines of Beringia during Glacial Maximum and Late Glacial times. But their full significance can be appreciated only in the broader context of data from all of Beringia, on the one hand, and from the more southern regions of our hemisphere, on the other. As such a perspective will become available only in the long term, it will suffice, at this time to discuss a few of the most pertinent archaeological issues.
Regional and Beringian implications
Over the 25-millennia span represented by the Bluefish Caves deposits, the few cultural signals that can be observed are clearly indicative of sporadic and generally diffuse manifestations belonging to the 15,000-year Pleistocene segment of the sequence. During this period, there are two sets of data that speak of more than just local human presence.
Firstly, there is the lithic series indicative of the presence of a microblade and burin industry. The tools in this series can be interpreted as elements of a regional facies of a technological complex that appears to be present throughout Beringia at the end of the Pleistocene (Note 12), and several examples of which survive during part of the Holocene. Vestiges of this complex have been uncovered at numerous sites in the Yukon and Alaska, where they form the core of what is subsumed by concepts such as the Denali complex (West 1967), the American Paleoarctic tradition (Anderson 1968; Dumond 1977) and the Beringian tradition (West 1981). This type of material is generally found together with technological forms such as bifaces, scrapers and blades. According to data from various sites in Alaska, particularly from the Nenana River region where it is currently being studied in detail, this techno-complex seems to have been well established in eastern Beringia by around 10,500 BP, close on the heels of the preceding Nenana complex. The latter, which dates from about 12,000 BP and is characterised by bifacial points and an absence of microblades, was recently described in broad terms, as being related to the Clovis tradition (sensu lato)(Powers and Hoffecker 1989).
This Nenana assemblage is considered by the above researchers as an expression, possibly late (Note 13), of the first human dispersals in eastern Beringia. In this context, the Bluefish data is more relevant to our perception of the chronology of cultural events than to their actual characterisation. Despite their meagreness and stratigraphic fuzziness, they suggest that bearers of an industry incorporating microblades and burins (and certainly more) had reached far-eastern Beringia well before the last millennium of the Late Glacial.
The same can be said for the other group of data, which in the Bluefish sequence corresponds to the beginning of the Glacial Maximum, between 23,000 and 25,000 BP. We are referring, in particular, to the mammoth bone objects (flake and core), produced by percussion, evidence that, in this instance, can be used to address issues of technological characterisation and chronology.
The context in which these objects were found at the Bluefish Caves enables us to reject outright a series of objections or counter-hypotheses (Note 14; Cinq-Mars and Morlan 1989) that have been recently put forward relative to the cultural interpretation given to similar bone remains found in the Old Crow lacustrine basin (Morlan 1980,1984). In fact, many of the bone specimens collected in the fossiliferous sites of the Old Crow basin bear attributes that are analogous in every respect to the essential elements of the “chaîne opératoire” described earlier. As the systematic repetition of such a series of actions cannot possibly be the result of environmental happenstance, we must consider their end-results as, if not tools themselves, then at the very least the products and by-products of cultural activities.
This evident relationship between the in situ Bluefish data and that obtained from the secondary deposits of the Old Crow Basin clearly indicates that our region, namely the Porcupine River Basin and the adjacent uplands, was the scene of a long series of cultural manifestations, the earliest of which, according to data from the Old Crow basin, seems to have occurred around 40,000 BP (Morlan, et al. 1990). In other words, even though examples have not yet been uncovered in Alaska and Siberia, it appears that the first detectable signs of a human presence in Beringia date back to the Wisconsinan interstadial.
Hemispheric implications
It follows from these findings that hypotheses concerning the antiquity of the first human exploration of our hemisphere, from its Beringian point of entry to Tierra del Fuego, should not be limited or influenced solely by the ecological, and therefore geographical constraints which characterised the Beringian Late Glacial and which are used, in conjunction with other factors, to support Clovis primacy in North America (Note 15).
Even if these constraints must be taken into consideration in an examination, for example, of the nature of the relationship between the Nenana complex and the Clovis cultures, they no longer hold up in an interstadial context. The “short chronology” mentioned above, could thus be viewed as one segment (among others) of a highly complex and much broader time-transgressive curve or trajectory that would serve to explain the archaeological manifestations found further south, and generally considered to be pre-Clovis or, to use Meltzer’s term (1989), “pre-12,000 BP” (Note 16).
Also, it may well be that our earliest data simply reflect an aborted colonisation event, which, for reasons unknown, never went any farther than the eastern Beringian threshold, and which bears no relationship to later manifestations. On the other hand, it is possible that this evidence is indicative of a long settling in, a pause during which there developed, in cultures as yet to be identified, modes of adaptation which subsequently would enable some of them to move south during the Late Glacial.
In conclusion, we would like to emphasise that the Bluefish data provide us with a clearer image of the time frame within which hypotheses dealing with the question of the initial peopling of America should be developed, and this for hypotheses that are strictly archaeological as well as for ones based on linguistic and biological data (Greenberg et al. 1986). Finally, these data make us realise that Beringia, as noted by Workman (1980), must be perceived not merely as a passageway between two continental poles, but as an important biological and cultural crucible of long duration, which should be studied as such if we are to eventually arrive at an understanding of the events that led to the peopling of America.
Significance of the Bluefish Caves in Beringian Prehistory – Page 5
BIBLIOGRAPHY
- Adovasio, J.M. and R.C. Carlisle.
1986 “Pennsylvania Pioneers”, Natural History Vol.95, pp. 21-27.
- Anderson, D.D.
1968 “A Stone Age Campsite at the Gateway to America”, Scientific American Vol. 218, pp. 24-33.
- Binford, L.R.
1981 Bones. Ancient Men and Modern Myths. (New York, Academic Press).
- Cinq-Mars J.
1979 “Bluefish Cave 1: A Late Pleistocene Eastern Beringian Cave Deposit in the Northern Yukon”, Canadian Journal of Archaeology No. 3, pp. 1-32.
- 1982 “Les grottes du Poisson-Bleu”, Geos Vol. 11, pp. 19-21.
- Cinq-Mars J. and B. Lauriol
1985 “Le karst de Tsi-it-toh-choh: notes préliminaires sur quelques phénomènes karstiques du Yukon septentrional, Canada, Annales de la Société Géoloqique de Belgique T. 108, pp. 185-195.
- Cinq-Mars, J. and R.E. Morlan.
1989 “The Peopling of the Americas as Seen from Northern Yukon Territory”, (article submitted for publication in the proceedings of the World Summit Conference on the Peopling of the New World, Orono, Maine, May 1989).
- Cinq-Mars, J. and E. Nelson.
1989 “AMS bone dates from the Bluefish Caves, northern Yukon, Canada”, (article under preparation, 1989).
- Cwynar, L. and J.C. Ritchie.
1980 “Arctic steppe-tundra: A Yukon perspective”, Science Vol. 208, pp. 1375-1377.
- Dillehay, T.D. and M. Collins.
1988 “Early Cultural Evidence from Monte Verde in Chile”, Nature Vol. 332, pp. 150-152.
- Dolitsky, A.B.
1985 “Siberian Paleolithic Archaeology: Approaches and Analytic Methods”, Current Anthropology Vol. 26, pp. 361-378. Dumond, D.E., 1977 The Eskimos and Aleuts. (London, Thames and Hudson).
- Fladmark, K.R.
1979 “Routes: Alternative Migration Corridors for Early Man in North America”, American Antiquity Vol. 44, pp. 55-69.
- 1982 “Microdebitage analysis: Initial Considerations”, Journal of Archaeological Science Vol. 9, pp. 205-220. Greenberg, J.H., C.G. Turner, II and S.L. Zegura.
- 1986 “The Settlement of the Americas: A Comparison of the Linguistic, Dental, and Genetic Evidence”, Current Anthropology Vol. 27, pp. 477-497.
- Gruhn, R.
1988 “Linguistic Evidence in Support of the Coastal Route of Earliest Entry into the New World”, Man (N.S.) Vol. 23, pp. 77-100.
- Gruhn, R. and A.L. Bryan.
1984 “The Record of Pleistocene Megafaunal Extinctions at Taima-Taima, Northern Venezuela”. In: Quaternary Extinctions: A Prehistoric Revolution, ed. P.S. Martin and R.G. Klein. (Tucson, University of Arizona Press) pp. 128-137.
- Guidon, N. and G. Delibrias.
1986 “Carbon-14 Dates Point to Man in the Americas 32,000 Years Ago”, Nature Vol. 321, pp. 769-771.
- Guthrie, R.D.
1982 “Mammals of the Mammoth Steppe as Paleoenvironmental Indicators. ” In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 307-326.
- 1985 “Woolly Arguments Against the Mammoth Steppe – A New Look at the Palynological Data”, Quarterly Review of Archaeology Vol. 6.
- Harington, C.R.
1980 “Radiocarbon Dates on Some Quaternary Mammals and Artifacts from Northern North America”, ArcticVol. 33, pp. 815-832.
- 1989 “Pleistocene Vertebrate Localities in the Yukon.” In: Late Cenozoic History of the Interior Basins of Alaska and the Yukon, ed. L.D. Carter, T.D. Hamilton and J.P. Galloway (U.S. Geological Survey Circular 10269) pp. 93-98.
- Haynes, C.V.
1982 “Were Clovis Progenitors in Beringia?” In: Paleoecology of Beringa, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 383-398.
- Hopkins, D.M.
1959 “Cenozoic History of the Bering Land Bridge (Alaska)”, Science Vol. 129, pp.1519-1528.
- Hopkins, D.M. (ed.)
1967 The Bering Land Bridge. (Stanford, Stanford University Press).
- Hopkins D.M., J.V. Matthews, Jr., C.E. Schweger and S. Young (eds.) 1982 Paleoecology of Beringia. (New York, Academic Press).
- Hughes O.L.
1972 “Surficial geology of northern Yukon Territory and northwestern District of Mackenzie”, Geological Survey of Canada Paper 69-36.
- Hultén, E.
1968 Flora of Alaska and neighboring territories. (Stanford University Press).
- Irving, W.N., N.A. Jopling and B. Beebe.
1986 “Indications of Pre-Sangamon Humans near Old Crow, Yukon, Canada.” In: New Evidence for the Pleistocene Peopling of the Americas, ed. A.L Bryan (Orono, Center for the Study of Early Man) pp. 49-63.
- Ives, J.W., A.B. Beaudoin and M.P.R. Magne.
1989 “Evaluating the Role of a Western Corridor in the Peopling of the Americas”, (communication prepared for the Circum-Pacific Prehistory Conference, Seattle, Washington, August 1989).
- Lauriol B., D.C. Ford and J. Cinq-Mars.
1989 “Landscape development from caves and speleothem data. Preliminary evidence from northern Yukon, Canada”, (article submitted to Journal canadien des sciences de la terre).
- Lumley, H. and M. de, M.C. Beltralo, Y. Yokoyama, J. Labeyrie, J. Danon, G. Delibrias, C. Falguères and J.L. Bischoff.
1987 “Présence d’outils taillés associés à une faune quaternaire datée du Pléistocène moyen dans la Toca da Esperanga, région de Central, état de Bahia, Brésil”, L’Anthropologie T. 91, pp.917-942.
- Lundelius, E.L., Jr.
1989 “The Implications of Disharmonious Assemblages for Pleistocene Extinctions”, Journal of Archaeoloaical Sciences Vol. 16, pp. 407-417.
- Martin, P.S.
1982 “The Pattern and Meaning of Holarctic Mammoth Extinction.” In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 399-408.
- Matthews, J.V., Jr.
1982 “East Beringia During Late Wisconsin Time: A Review of the Biotic Evidence.” In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 127-150.
- Mochanov, Yu.
1978 “The Paleolithic of NE Asia and the problem of the first peopling of America.” In: Early Man in America from a Circum-Pacific perspective, ed. A.L. Bryan (University of Alberta Department of Anthropology Occasional Paper 1) p. 67.
- 1980 “Early migrations to America in the light of a study of the Dyuktai Paleolithic culture in Northeast Asia.” In: Early Native Americans, ed. D.L. Brownman (La Haye, Mouton) pp. 119-131.
- Morlan, R.E.
1980 Taphonomy and archaeology in the Upper Pleistocene of the northern Yukon Territory: a glimpse of the peopling of the New World. (Ottawa, National Museum of Man, Mercury Series, Archaeological Survey of Canada, paper no. 94).
- Morlan, R.E.
1984 “Toward the Definition of Criteria for the Recognition of Artificial Bone Alterations.” Quaternary Research Vol. 22, pp. 160-171.
- Morlan, R.E.
1989 “Paleoecological implications of Late Pleistocene and Holocene microtine rodents from the Bluefish Caves, northern Yukon Territory”, Canadian Journal of Earth Sciences Vol. 26, pp. 149-156.
- Morlan, R.E. and J. Cinq-Mars.
1982 “Ancient Beringians: Human Occupation in the Late Pleistocene of Alaska and the Yukon Territory.” In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 353-381.
- Morlan, R.E., D.E. Nelson, T.A. Brown, J.S. Vogel and J.R. Southon.
1989 “Accelerator Mass Spectrometry Dates on Bones from the Old Crow Basin”, Canadian Journal of Archaeology Vol. 14, pp. 75-92.
- Norris, D.K.
1985 Stratigraphy of Devonian Outcrop Belts in Northern Yukon Territory and Northwestern District of Mackenzie (operation Porcupine area). (Ottawa, Bulletin 410, Geological Survey of Canada).
- Powers, W.R. and J.F. Hoffecker.
1989 “Late Pleistocene Settlement in the Nenana Valley, Central Alaska”, American Antiquity Vol. 54, pp. 263-287.
- Ritchie, J.C.
1984 Past and Present Vegetation of the Far Northwest of Canada. (Toronto, University of Toronto Press).
- Ritchie J.C., J. Cinq-Mars and L. Cwynar.
1982 “L’environnement tardiglaciaire du Yukon septentrional, Canada”, Géographie physique et Quaternaire Vol. XXXVI, pp. 241-250.
- Ritchie, J.C. and L. Cwynar.
1982 “The Late Quaternary Vegetation of the North Yukon.” In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 113-126.
- Roberge, J., B. Lauriol, P. Thibaudeau and J. Cinq-Mars.
1986 “Caractère des karsts arctiques du Yukon septentrional”, Comptes rendus du IXème Congrès international de spéléologie (Barcelone), Vol. 1, pp. 164-167.
- Schweger, C.E., J.V. Mathews, Jr., D.M. Hopkins and S.B. Young (eds.)
1982 “Paleoecology of Beringia – A Synthesis.” In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 425-444.
- Shipman, P.
1981 “Applications of Scanning Electron Microscopy to Taphonomic Problems. ” In: The Research Potential of Anthropological Museum Collections, ed. A.-M. Cantwell, J. Griffin and N.A. Rothschild (New York, Annals of the New York Academy of Sciences 376), pp. 357-386.
- Vereshchagin, N.K. and O.F. Baryshnikov.
1982 “Paleoecology of the Mammoth Fauna in the Eurasian Arctic.” In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press), pp. 267-279.
- West, F.H.
1967 “The Donnelly Ridge Site and the Definition of an Early Core and Blade Complex in Central Alaska”, American Antiquity Vol. 32, pp. 300-382.
- West, F.H.
1981 The archaeology of Beringia. (New York, Columbia University Press).
- Workman, W.B.
1980 “Holocene Peopling of the New World: Implications of the Arctic and Subarctic Data.” In: The Ice-Free Corridor and Peopling of the New World, ed. N.W. Rutter and C.E. Schweger (Canadian Review of Anthropology, Vol. 1), pp. 129-139.
- Yi, S. and G. Clark.
1985 “The ‘Dyuktai Culture’ and New World Origins”, Current Anthropology Vol. 26, pp. 1-20.
Significance of the Bluefish Caves in Beringian Prehistory – Page 6
NOTES
- We refer here to Beringian options in the strict sense and to the concept of the “Mackenzie corridor” (Ives et al. 1989), without denying the value of the “coastal” hypothesis formulated by Fladmark (1979) and taken up by Gruhn (1988).
- The caves were discovered accidentally in 1975, and sampling and excavations were conducted intermittently between 1977 and 1987 (Cinq-Mars 1979 and 1982; Morlan and Cinq-Mars 1982).
- As analysis has demonstrated that the loess is not from the surrounding limestone landscape (C. Tarnocai, pers. comm.), we must seek its origin in sediments that became available following the drainage of the lake basins to the north (Bluefish and Old Crow glacial lakes).
- Results of a study of part of the microtine assemblages from Caves I and II, recently conducted by R. Morlan (1989), provide another example of this contrast.
- All the dates of faunal specimens were run on bone collagen and, but for two exceptions, were obtained by accelerator mass spectrometry (AMS), thanks to R.M. Brown (Atomic Energy of Canada) and E. Nelson (Simon Fraser University).
- The controversy concerns the precise nature and the relative viability of the Beringian environment during the Last Glacial Maximum. The whole question is known as that of the “productivity paradox” (Schweger et al. 1982). Some (Cwynar and Ritchie 1980) assert that these regions could have supported, at best, a greatly impoverished biome, while others contend that the environment was extremely rich (Guthrie 1985; Matthews 1982).
- Two objects (a core and a microblade fragment) were discovered in the humus-rich rubble of Cave II, in a highly disturbed context. Surface finds of small chips and a microblade were also made on top of one of the ridges of the Bluefish limestone outcrop.
- Bluefish limestone does not contain chert. The few examples of this material found in the region (in several rocky outcrops to the east of the caves or in the form of pebbles on the river bars) are of very poor quality. We also know that cherts similar to those found in Caves I and II occur in a number of archaeological sites located about 100 kilometres to the north, in the Cordilleran foothills (Brooks, British and Barn mountains).
- These flakes, generally smaller than 0.003 m, were collected during the processing of sediment samples (Cinq-Mars 1979; Fladmark 1982).
- To date, these cut marks, which are often difficult to read, have been examined with a binocular microscope. We plan in the near future to confirm our findings with the help of scanning electron microscopy (SEM), following the procedure suggested by Shipman (1981).
- This flake also belongs to the previous category, as the dorsal face exhibits a short cut mark.
- This technological complex is better known in Siberia as the Dyuktai culture or tradition (Mochanov 1978, 1980). Although it would not be appropriate here to discuss its origins, age and significance, we should mention that these issues are far from being resolved (Dolitsky 1985; Yi and Clark 1985). Nevertheless, according to these authors, there appears to be clear evidence of its presence in northeastern Siberia at the end of the Glacial Maximum, around 18,000 BP. This is in agreement with the age that we would assign to it in eastern Beringia, on the basis of the Bluefish data.
- The location of the cultural manifestations studied by Powers and Hoffecker (1989) can perhaps be explained by the fact that at the end of the Late Glacial, the Nenana valley and several neighbouring regions served as refugia where survived the last traces of the herbaceous tundra and Mammoth Steppe fauna. It can be assumed that the human groups who were living there were carrying on with modes of adaptation which were probably pan-Beringian, and of necessity, derived from earlier times.
- The reader can find a critique (Cinq-Mars and Morlan 1989) of the counter-hypotheses which view the percussion-flaked mammoth bones found in the fossiliferous deposits of the Old Crow River as having been shaped by various natural phenomena. We do not, however, altogether deny the validity of these counter-hypotheses as possible explanations for several other types of modifications found on the bones of various other taxa.
- We refer here to the theses put forward by Paul Martin (1982) and Vance Haynes (1982).
- Without necessarily accepting them outright, we refer to sites such as Meadowcroft (Adovasio and Carlisle 1986), Monte Verde (Dillehay and Collins 1988), Toca do Boqueirao da Pedra Furada (Guidon and Delibrias 1986) and Taima-Taima (Gruhn and Bryan 1984). On the other hand, it is clear that the Beringian chronological frame of reference that we have tried to illustrate, unlike that proposed by Irving and his colleagues (Irving et al. 1986), still cannot accommodate a site such as Toca da Esperança (de Lumley et al. 1987).